
Like the blind men and the elephant, perspective drives understanding in ecology. The temporal and spatial scale of analysis (let alone the system and species you focus on) has major implications for your conclusions. Most ecologists recognize this fact, but consider only particular systems, scales or contexts due to practical limitations (funding, reasonable experimental time frames, studentship lengths).
Ecologists have long known that regional processes affect local communities and that local processes affect regional patterns. Entire subfields like landscape ecology, metapopulations, metacommunities, and biogeography (species area relationships) highlight these spatial dependencies. But high-profile ecological research into biodiversity and ecosystem functioning ('BEF') primarily considers only local communities. Recently though, the literature has started to fill this gap and asking what BEF relationships look like at larger spatial scales, and how well local BEF relationships predict those at larger spatial scales.
'Traditional' BEF experiments were done at relatively small spatial scales (often only a few meters^2). Positive BEF relationships were commonly observed, but often were quite saturating – that is, only a few species were necessary to optimize the function of interest. If the impact of biodiversity saturates with only a few species, it would seem that surprisingly few species are necessary to maintain functioning. True, studies that considered multiple ecosystem functions are more likely to conclude that additional diversity is required for optimal functioning (e.g. Zavaleta et al. 2010). But a simplistic evaluation of the facts that a) ecosystem functioning rapidly saturates with diversity, and b) locally, diversity may not be generally decreasing (Vellend et al. 2017), could lead to overly confident conclusions about the dangers of biodiversity loss. Research on BEF relationships, as they transition from local to larger spatial scales, is increasingly suggesting that our understanding is incomplete, and that BEF relationships can grow stronger at large spatial scales.
A number of recent papers have explored this question, and in considering the essential role of spatial scale. Predictions about how BEF relationships will change with spatial scale vary. On one hand, in most systems there are only a few dominant species and these species may disproportionately contribute to ecosystem functions, regardless of the spatial scale. On the other hand, species-area relationships tend to increase rapidly at small scales, as community composition turns over. If that is the case, then different species may make important contributions in different places. Winifree et al. (2018) contrasted these predictions for three crop species that rely on natural bee pollinators (cranberries, blueberries, and watermelons). They censused pollinators at 48 sites, over a total extent of ~3700 km^2. Though at local scales very few bee species were required to reach pollination goals, the same goals at larger spatial scales required nearly an order of magnitude more bee species. These results in particular appeared to be driven by species turnover among sites--perhaps due to underlying environmental heterogeneity.
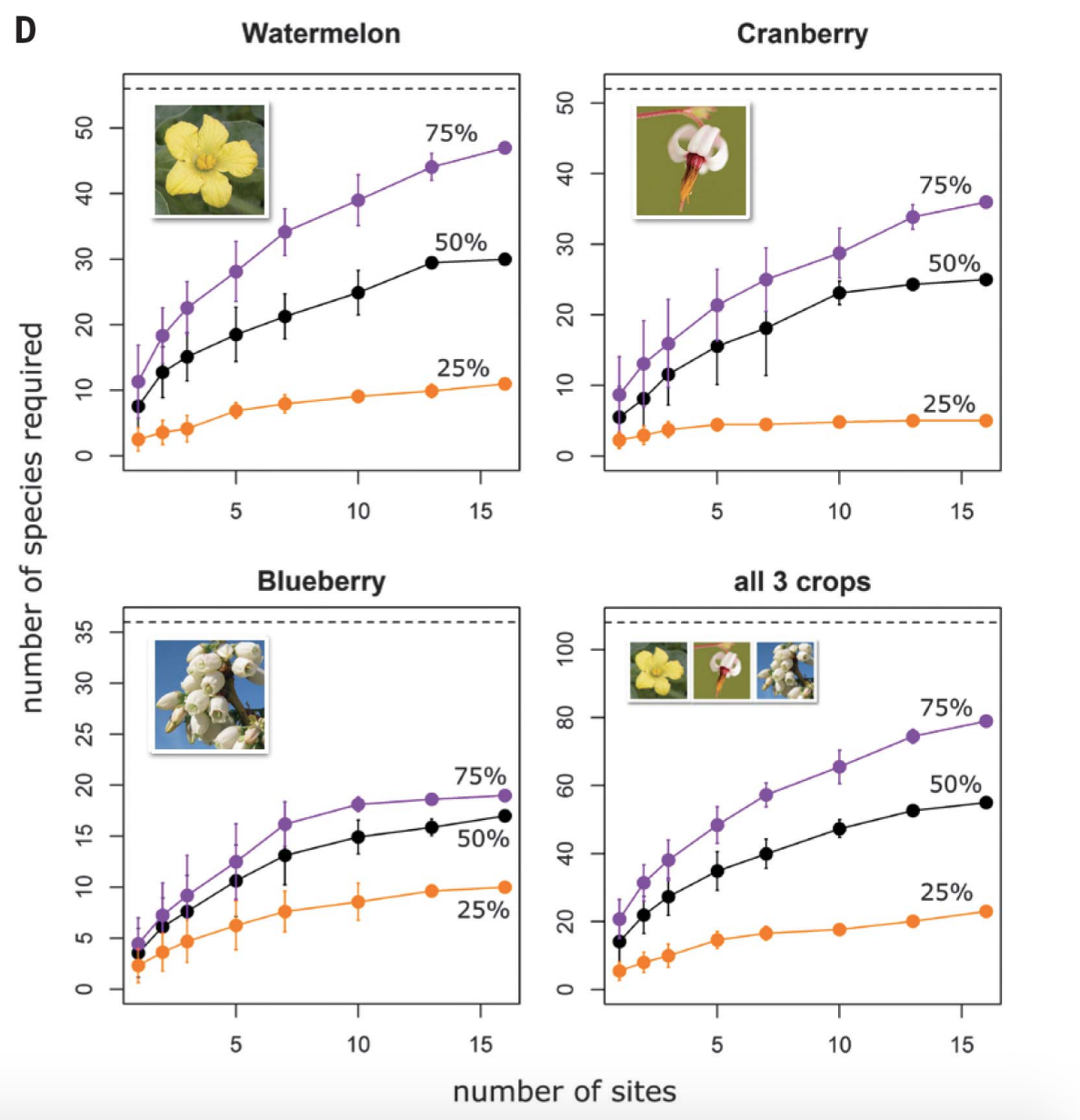 |
| From Winifree et al. "Cumulative number of bee species required to maintain thresholds of 25% (orange), 50% (black), and 75% (purple) of the mean observed level of pollination, at each of n sites (16). Horizontal dashed lines indicate the total number of bee species observed in each study. Error bars represent 1 SD over all possible starting sites for expanding the spatial extent. For all three crops combined, each x-axis increment represents the addition of one site per crop". |
Another mechanism for increased BEF at larger scales is insurance effects. The presence of greater diversity can interact with spatial and temporal environmental variation to increase or stabilize ecosystem functioning. Greater diversity should maximize the differential responses of species to changing conditions, and so buffer variation in ecosystem functioning. Such effects, when they occur through time are temporal insurance, and when they occur via dispersal among sites, spatial insurance. Wilcox et al. (2018) considered the role of synchrony and asynchrony among populations, communities, and metacommunities to ask whether local asynchrony affected stability (see Figure below for a nice conceptual explanation). Across hundreds of plant data sets, they found that asynchrony of populations did enhance stability. However, the degree to which it affected stability varied from very weak to very important (e.g. by 1% to 300%). Maximizing species or population differences at local scales apparently can have implications for dynamics, and so potentially stability of functioning, at much larger scales.
Finally, Isbell et al. (2018) describe ways in which ecosystem functioning and other contributions of nature to humanity are scale-dependent, laying out the most useful paths for future work (see figure below).
These papers make nearly identical points worth reiterating here: 1) we have done far too little work beyond the smallest spatial scales (~3 m^2) and so lack necessary knowledge of the impacts of losing of biodiversity, and 2) policy decisions and conservation activities are occurring at much larger scales – at the scale of the park, the state, or the nation. Bridging this gap is essential if we are to make any reasonable arguments as to why ecosystem function figure into large-scale conservation activities.
Plant biodiversity change across scales during the Anthropocene. Vellend, Mark, et al. Annual review of plant biology 68 (2017): 563-586.
Species turnover promotes the importance of bee diversity for crop pollination at regional scales. RACHAEL WINFREE, JAMES R. REILLY, IGNASI BARTOMEUS, DANIEL P. CARIVEAU, NEAL M. WILLIAMS, JASON GIBBS. SCIENCE16 FEB 2018 : 791-793
Asynchrony among local communities stabilises ecosystem function of metacommunities. Kevin R. Wilcox, et al. Ecology Letters. Volume 20, Issue 12, Pages 1534–1545.
Isbell, Forest, et al. "Linking the influence and dependence of people on biodiversity across scales." Nature 546.7656 (2017): 65.
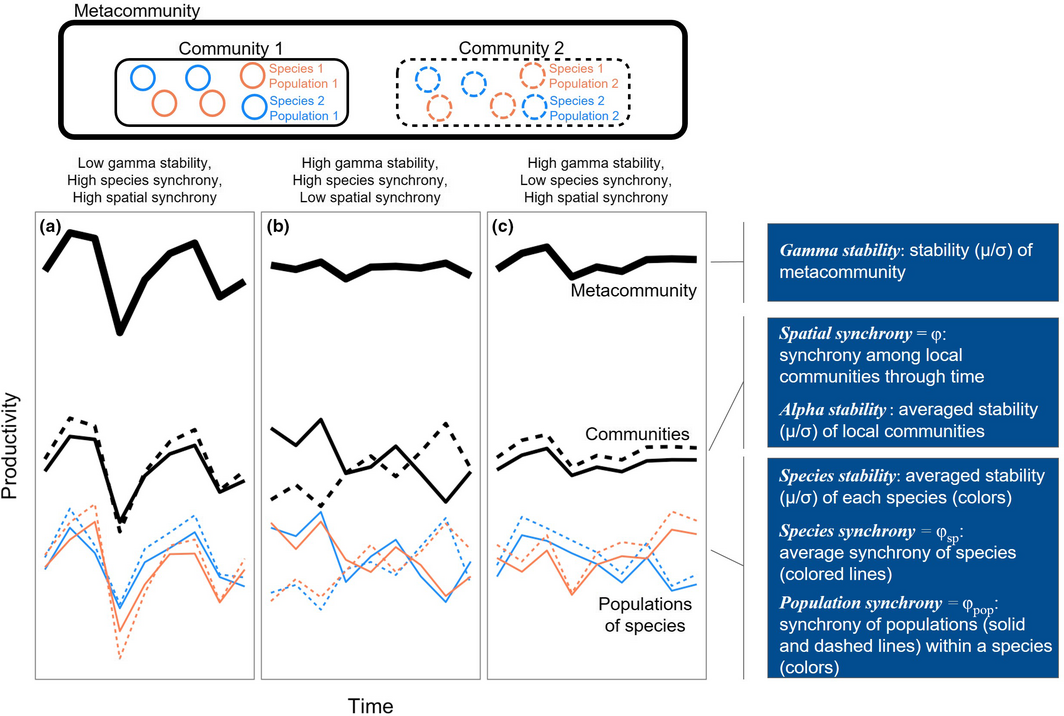 |
| From Wilcox et al. "Conceptual figure showing how stability and synchrony at various spatial scales within a metacommunity combine to determine the stability of ecosystem function (here, productivity). In (a), high synchrony of species within and among local communities results in low stability at the scale of the metacommunity. In (b), species remain synchronised within local communities, but the two communities exhibit asynchronous dynamics due to low population synchrony among local patches. This results in relatively high gamma stability. Lastly, in (c), species exhibit asynchronous dynamics within local communities through time, and species-level dynamics are similar across communities (i.e. high population synchrony). This results in relatively high gamma stability. Blue boxes on the right outline stability components and mechanisms, and the hierarchical level at which they operate. Adapted from Mellin et al. (2014)." |
 |
| From Isbell et al. 2018. |
References:
Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Erika S. Zavaleta, Jae R. Pasari, Kristin B. Hulvey, G. David Tilman. Proceedings of the National Academy of Sciences Jan 2010, 107 (4) 1443-1446; DOI: 10.1073/pnas.0906829107.
Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Erika S. Zavaleta, Jae R. Pasari, Kristin B. Hulvey, G. David Tilman. Proceedings of the National Academy of Sciences Jan 2010, 107 (4) 1443-1446; DOI: 10.1073/pnas.0906829107.
Species turnover promotes the importance of bee diversity for crop pollination at regional scales. RACHAEL WINFREE, JAMES R. REILLY, IGNASI BARTOMEUS, DANIEL P. CARIVEAU, NEAL M. WILLIAMS, JASON GIBBS. SCIENCE16 FEB 2018 : 791-793
Asynchrony among local communities stabilises ecosystem function of metacommunities. Kevin R. Wilcox, et al. Ecology Letters. Volume 20, Issue 12, Pages 1534–1545.
Isbell, Forest, et al. "Linking the influence and dependence of people on biodiversity across scales." Nature 546.7656 (2017): 65.