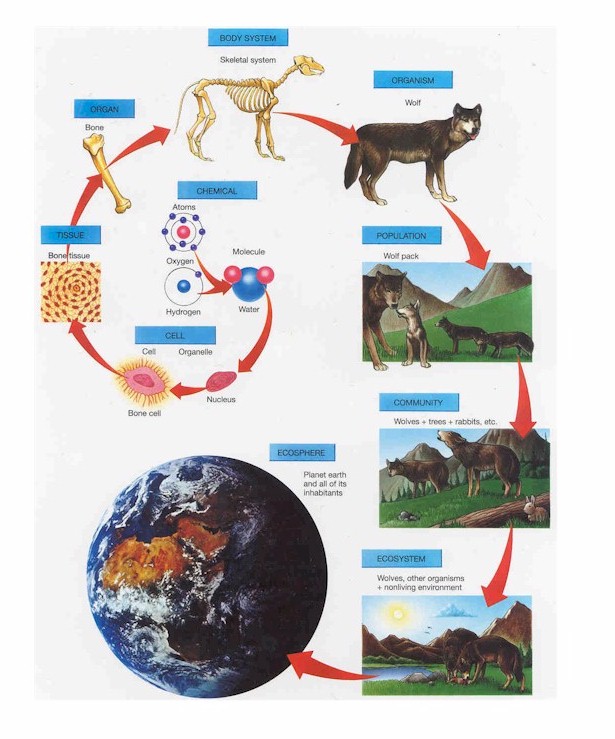
Particularly, the functional ecology literature has developed increasingly complex and integrative methods for measuring and comparing trait diversity. The literature has gone from descriptions of general types or traits (e.g. Whittaker 1956), to measuring measuring individual traits and relating them to particular ecologically relevant variables (e.g. Gaudet and Keddy (1988)); to calculating community-weighted values for individual traits (e.g. D Schluter, (1986)); to incorporating multiple variables into single measures (e.g. FD package); to a framework reflecting mathematical moments in data (Villeger et al. (2008); and to the use of multivariate hypervolumes to describe the multi-dimensional shape and volume of trait space to be measured (e.g. Blonder et al. 2014).
A new paper in TREE does a nice job of summarizing and integrating these developments with yet another addition: a ‘trait probability density’ approach. In “Traits Without Borders: Integrating Functional Diversity Across Scales", Carlos P. Carmona, Francesco de Bello, Norman W.H. Mason, and Jan Lepš nicely illustrate a way to capture the complexity inherent to a concept such as the ‘functional niche’. [The "region of the functional space containing all the trait combinations displayed by the individuals of a species"].
The truth about traits is that there is meaningful variation at every scale at which we measure them (including variation between individuals, variation between populations, variation between species, and variation between communities). Often decisions are made to ignore or collapse unwanted levels of variation (such as using a mean value across several individuals to calculate a single species-level value). The authors suggest that we can instead incorporate this variation appropriately. A probability density function can be defined for the multi-trait space, with probabilities representing the relative abundances of each combination of trait values. Thus, for a species, the curve (Figure IA) would show the multivariate trait space seen across all measured individuals, with uncommon combinations of traits seen in few individuals shown at the tails of the distribution. Outliers and extreme values are incorporated but not overemphasized as they can be in convex hull approaches.
The probabilistic approach reflects that a niche *is* probabilistic for a species - after all, it is unlikely that the niche is simply a fixed set of traits that is identical for all individuals or populations. However, not all combinations of trait values (niche dimensions) are equally likely for members of a species, and these curves reflect that. And when probabilities are incorporated into trait measurements, greatly different conclusions may be made about how similar or dissimilar assemblages may be (e.g. Fig IC).
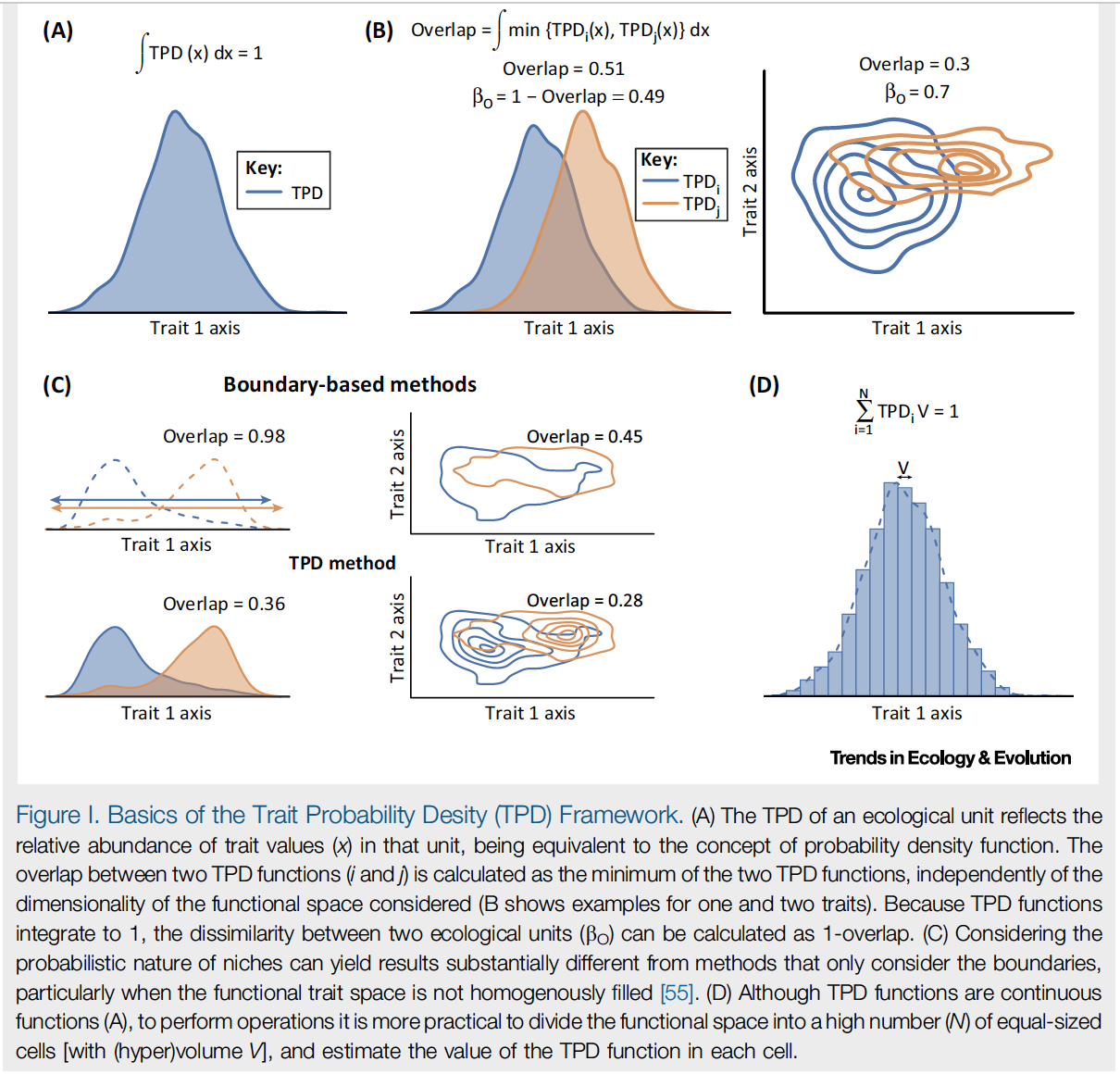 |
| Reproduced from Fig I., Carmona et al. 2016 TREE. |
One concern--one that is pretty much universal to all analyses in functional ecology--is about how the biases and limitations of available data will affect this type of measure. Some species are better described, some traits are not available for most species, some studies lack interspecific measures, some lack local measures (relying instead on general databases of trait values). In addition, some intraspecific variation arises from other sources of noise like stochasticity and measurement error. This is all part of a bigger question about sufficient data: not only do we need to know how many traits are needed to define a species, but we need to decide how much and what kind of data is necessary to understand a trait…
 |
| Fig. 2 from Carmona et al. TREE 2016. It is possible to incorporate existing measures of functional diversity (richness, evenness, divergence) into the probabilistic definition. |