www.pixshark.com
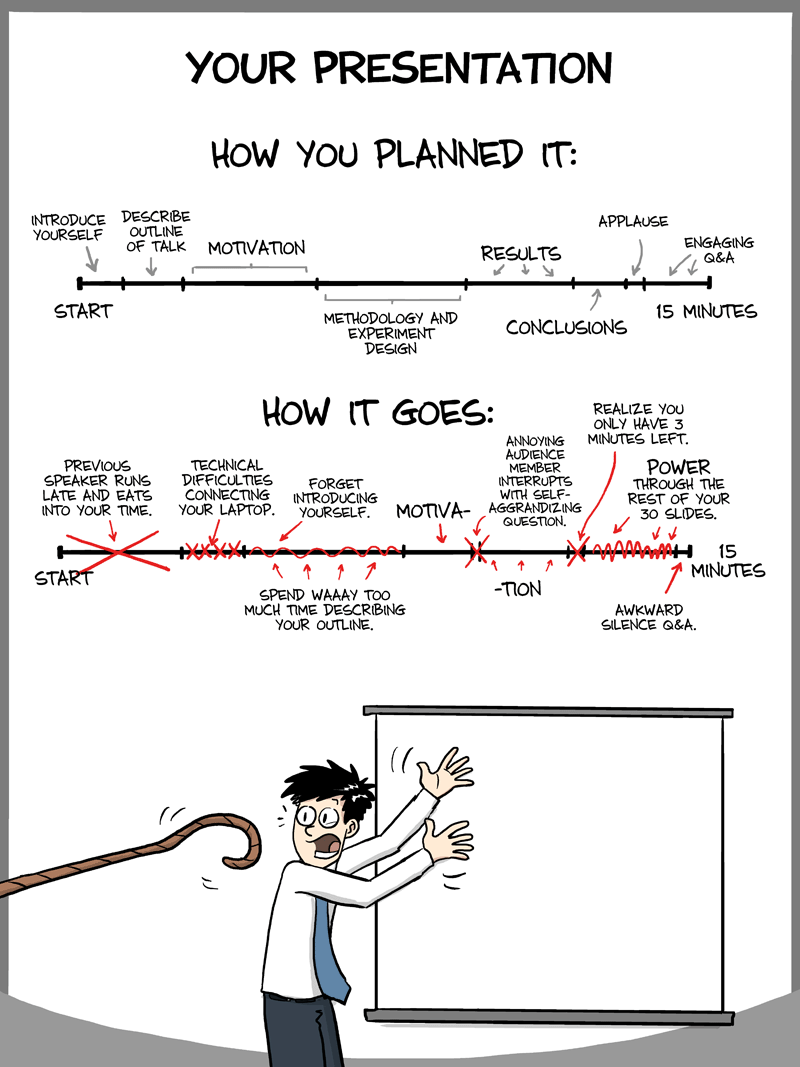
With the ESA submission deadline just passing, the Cadotte
Lab decided that it would be helpful to dish out a few tips on how to make a
presentation that is both enjoyable for your audience and fun for you to give. Presenting
in front of people is never easy; giving a presentation about your own study
can be even harder since you have to condense months (or even years) worth of
information into a 15 minute time period. So with this in mind here are a few
tips for each of the main sections of a presentation:
Note, the percentage by each section heading indicates the
relative amount of time you should spend on that section.
Title Slide (5%)

www.nichecartoons.com
This is the first chance you’ll get to catch your audience’s
attention, so be interesting!
The title of your presentation depends on the type of
audience you’ll be presenting to, so gauge it accordingly. If your audience is
a bunch of people with only general biology backgrounds or people that are from
completely different fields then don’t complicate things using heavy jargon.
Generally for the title, you want to:
- Be witty and interesting
- Convey the main message or main result from your study
If you’re speaking to a broad audience it could be helpful
to have a broad title and then separate it from a more specific title.
Besides the title you’ll also want to include your name and
affiliation. Depending on the type of talk, for instance an honors thesis, you
should also include your supervisor’s name. If you are collaborating with many
people on a study you should also include their names. However, make sure that
your name is on the first slide, since you are the presenter, and then on a
second slide include a special acknowledgement of the other people involved.
It’s also recommended that you acknowledge these people throughout the talk,
such as in the methods.
Introduction (10-15%)
Don’t make this section too long. Give just enough
background that the audience can understand the concepts that you’ll be
discussing and how it relates to the question you are trying to answer.
Generally for the introduction, you want to:
- Have the background information displayed in a simple to understand way
- You could use info-graphs here to reinforce an idea
- By the 2nd or 3rd slide you’ll want to state your study objectives or hypotheses
- You could create ‘toy’ graphs to describe your hypotheses / predictions
Methods (10-15%)
Be very concise with this section. Everyone understands that
a lot of work went into performing your study; however, you don’t want to
overwhelm your audience with all the nitty-gritty things you had to do. Give
enough detail that people understand what you did and if possible try and
summarize your methods in a simple figure.
Generally for the methods, you want to talk about:
- The treatments used, sample size, the measurements taken and how they were done, and the statistics that you performed
A note on statistics: try to steer clear of very complicated
statistics. Most likely your audience will have a basic understanding of stats,
but you may lose people if you get too complicated. When talking about your
stats, make sure that you can give an easy to understand explanation of how
they work.
Results (50%)
This is the biggest and best section; it’s where you get to
show people all the cool and exciting things you’ve done! However, the only way
you can convey how awesome your results are is by clearly explaining them.
Generally for the results, you want to:
- Stick to the main results
- You may have a lot different results but always make sure that what you are describing relates directly to the main message of your study
- Don’t overwhelm your audience
- Always thoroughly describe your graphs
- Describe what variables were you examining (the axes)
- Why is the graph important?
- What is the relationship that the graph is showing?
- The title of the slide could be used to state what the result is
- You’ve spent a lot of time making these graphs and analyzing them - so you know them very well, but your audience doesn’t yet. Take time to walk them through the graphs.
- If you’re showing several graphs in sequence, make sure to note if the axes are changing
- If the graphs are very similar it might be helpful to have a break between slides or to use an animation.
- Don’t show too many stats
- Just state the p-values and which stats were used
- Avoid tables if possible
- Summarize all the information in an easy to follow figure
- If you can’t avoid using a table make it as appealing as possible
- Highlight key parts or add arrows to show trends if they exist
Discussion (20%)
Now start bringing everything back together. Your audience
may have gotten lost during your results section, so now is the time to refocus
them so that they can see the big picture.
Generally for the discussion, you want to:
- Restate your hypotheses
- Restate you main results
- Describe how you could improve your study
- Describe the next steps for your work and the field in general
In the end you’ll want to describe the broader implications
of your work and give the audience a take home message so that they know that
your work is bettering the field in some way.
Acknowledgements (5%)
Don’t forget to thank everyone who has helped you through
this whole process! This includes your supervisor, people who helped you with
data analysis or revising your paper, or all the volunteers you helped you
conduct your field work or lab work. You’ll also want to acknowledge your
institution as well as anyone who provided funding to your project.
General tips
Here’s a quick list of tips to use throughout your presentation:
- Use large text font
- Don’t be flashy, make sure it’s easy to read
- Don’t put too much text on a slide
- This distracts the audience
- Don’t put any important point (text or an image) at the bottom 1/3rd of a slide
- Depending on the room you are presenting in it may be very hard for the audience to see it
- In general, try and keep everything within the top 2/3rd of the slide
- Don’t put too many animations on a slide
- This can be very distracting for the audience
- Don’t read off your slides
- Use presenter view if you can’t memorize everything
- Including outlines
- Not necessary in a short talk, but could be helpful in a longer talk
- If you run out of time
- Panic on the inside not the outside!
- Acknowledge that you’re running out of time and start wrapping things up
- Start talking about the broad implications of your work and maybe future directions you plan to take
- If you have more slides, skip over them but tell the audience what you were planning on showing. If they ask questions about what you were going to show you can go back to those slides
- Don’t talk too fast!
- Everyone gets nervous! Take a deep breath and calm yourself down, the calmer you are the easier it is for your audience to follow you