Community ecologists talk about communities as experiencing particular processes in a rather general way. We fall into rather Clementsian language, asking whether environmental filtering dominates a community or if biotic interactions are disproportionately strong. This is in contrast to the typical theoretical focus on pairwise interactions, as it acts as though all species in a community are responding similarly to similar processes.
Some approaches to community ecology have eschewed this generality, particularly those that focus on ecological ‘strategies’ differentiating between species. For example, Grimes argued that species in a community represented a tradeoff between three potential strategies - competitive, stress-tolerant, and ruderal (CRS).
Other related work describes rarity as the outcome of very strong density-dependence. The core-transient approach to understanding communities differentiates between core species, which have deterministic dynamics tied to the mean local environment, in contrast to transient species which are decoupled from local environmental conditions and have dynamics are driven by stochastic events (immigration, environmental fluctuations, source-sink dynamics). Assuming environmental stationarity, core species will have predictable and consistent abundances through time, in comparison to transient species.
If species do respond differently to different processes, then attempting to analyse all members of a community in the same way and in relation to the same processes will be less informative. Tests for environment-trait relationships to understand community composition will be weaker, since the species present in a community do not equally reflect the environmental conditions. In “
A core-transient framework for trait-based community ecology: an example from a tropical tree seedling community”, Umana et al (2017) ask whether differentiating between core and transient species can improve trait-based analyses. They analyse tropical forest communities in Yunnan, China, predicting that core species "will have strong trait–environment relationships that increase the growth rates and probability of survival that will lead to greater reproductive success, population persistence and abundance".
The data for this test came from 218 1 m2 seedling plots, which differed in soil and light availability. The authors estimated the performance of individual seedlings in terms of relative growth rate (RGR). They also gathered eight traits related to biomass accumulation, and stem, root and leaf organ characteristics. They were particularly interested in how the RGR of any individual seedling differed from the mean expectation for their species. Did this RGR deviation relate to environmental differences between sites? If a species’ presence is strongly influenced by the environment, then RGR deviation should vary predictably based on environmental conditions.
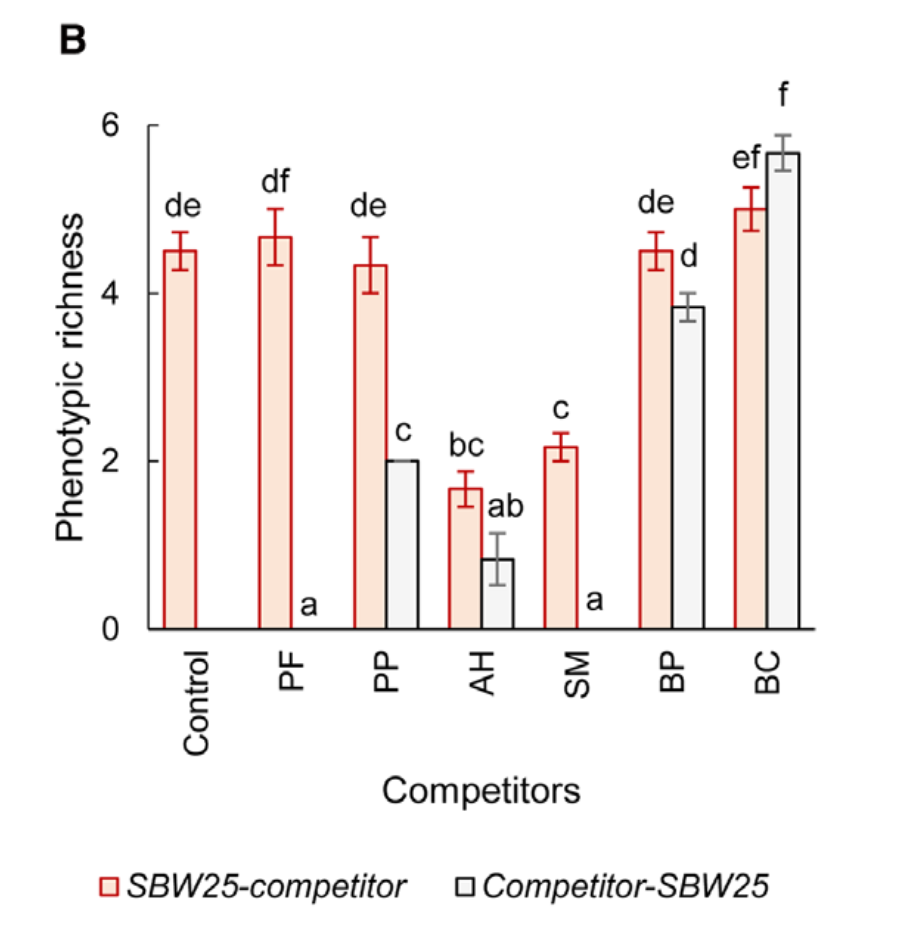
They then modelled RGR deviation as a function of the traits or environmental conditions (PCA axes). They considered various approaches for binning species based on commonness vs. rarity, but the general result was that bins containing rarer species had fewer PCA axes significantly associated with their RGR deviation and/or those relationships were weaker (e.g. see Figure below).
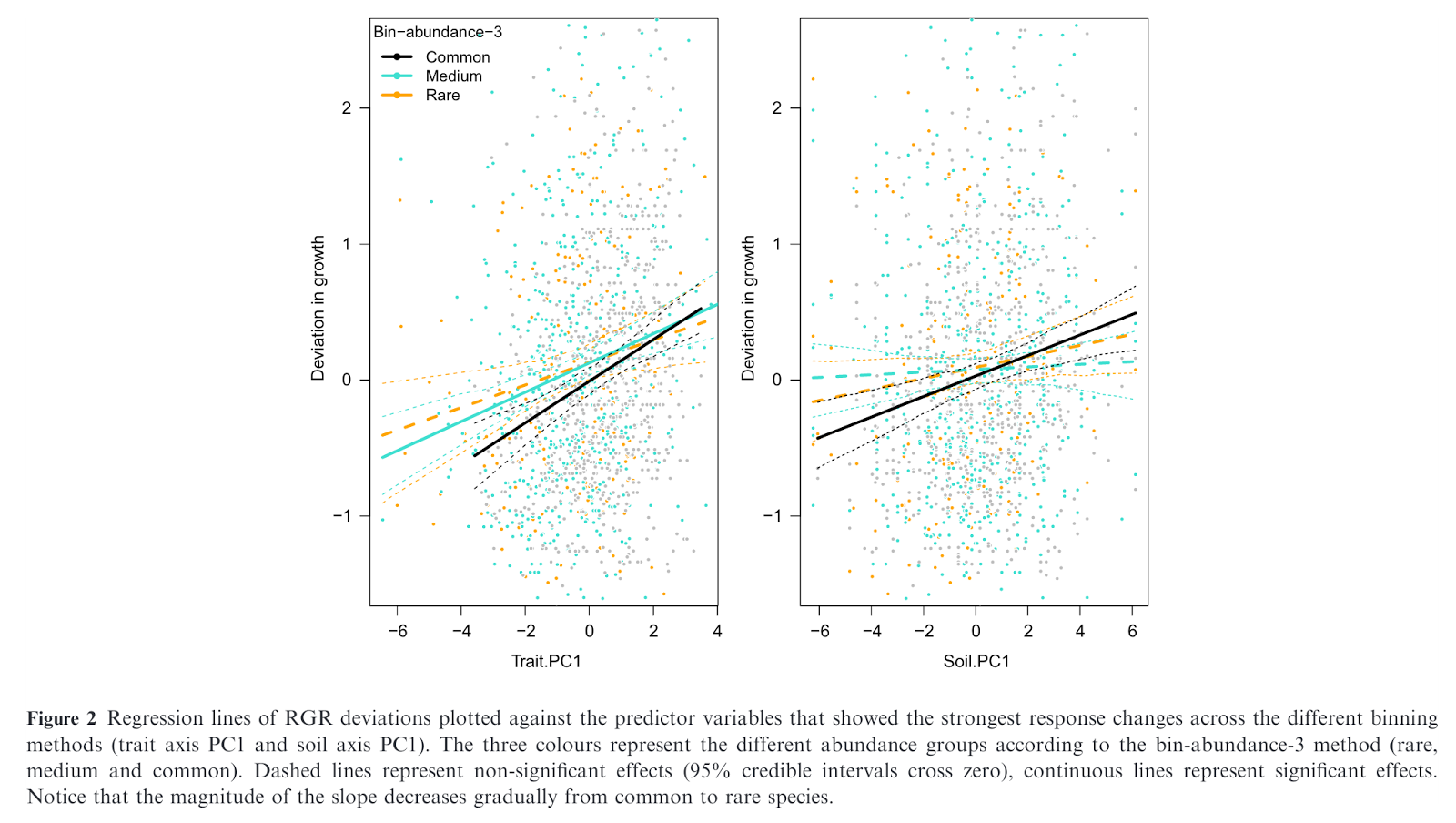
They conclude that “the main results of our study show that the strength of demography-environment/trait and trait-environment relationships is not consistent across species in a community and the strength of these effects is related to abundance”. Note that other studies similarly find variation in the apparent mechanism of coexistence in communities. For example,
Kraft et al. 2015 found that local fitness and niche differences only predict coexistence for a fraction of species co-occurring in their sites.
Umana et al.'s result is a reminder that work looking for general processes at the community level may be misleading. It isn't clear that there is a good reason to divide species into only two categories (e.g. core versus transients): like unhappy families, transient species may each be transient in their own way.