I was recently asked “what is the most important problem in ecology?”. I was dissatisfied with whatever I ended up answering, so it has been on my mind. I think there is an analogy with medicine here – it’s a little like asking a medical scientist “what is the most important disease to cure?” Similarly, there are multiple possible answers, and the one you give will depend on your area of interest/what type of doctor you are. (I also assume this is a question about basic research, and the answer is not as simple as saying, stop extinctions or prevent habitat loss).
 |
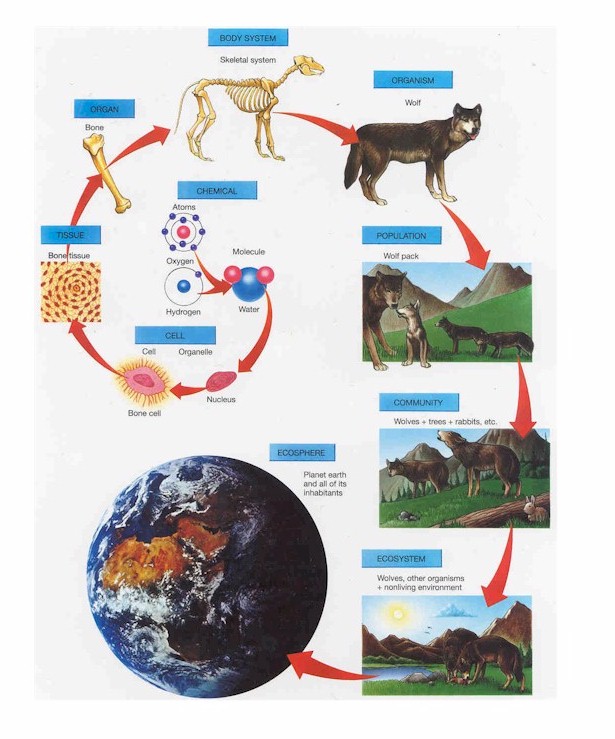
| Levels of biological organisation. |
The medical analogy breaks down a little because ecology is *far* more complicated than medical science. Medicine has a foundation in anatomy and physiology, which in turn rely on basic sciences like cell biology and genetics. This creates a reasonably constrained framework within which further learning/investigation can be organized. Medicine typically stops at the level of the individual, but ecology inherently involves many additional levels of organization (from individuals, to populations, to species and communities, to ecosystems, and beyond). Within any one of these higher levels of organization (population, community, ecosystem), there can be such an immense amount of variation in outcomes and dynamics that ecologists can lose sight of connections with lower and higher levels. For example, community ecology encompasses so much complexity on its own, that also considering the impacts of population level processes and on ecosystem level processes is a tall order. But, we should also appreciate, given these barriers to understanding, just how far ecology has actually advanced in the last 100 years. The combination of reductionist experiments and descriptive work at all scales has been immensely successful (e.g. see
this blog post for a partial list). Many general tools have been developed that we can then use to answer specific ecological questions (the integration with statistics with ecology h
as been highly successful; the use of specific mathematical models). Still, the ability to reconcile multiple levels of organization and scales still limits ecology.
This is a problem that cell biology has also experienced, and is now approaching via systems biology: "The reductionist approach has successfully identified most of the components and many of the interactions but, unfortunately, offers no convincing concepts or methods to understand how system properties emerge...the pluralism of causes and effects in biological networks is better addressed by observing, through quantitative measures, multiple components simultaneously and by rigorous data integration with mathematical models"(1): to me, this quote rings so true for ecology as well. Systems biology uses mechanistic, mathematical and computational models to attempt to represent multi-scale complexity.
Of course, the optimism about systems biology might be premature in that it hasn’t produced many useful models yet, such that it may be
“more of an agenda than a body of results.”. Some of the best “systems ecology” (e.g. meta-ecosystem models) are very system specific and data-heavy (e.g. 2). Can they inform us about generality in ecology?
All of which is to say, I think the most important problems in ecology relate to this need to make the connections between studies and systems and levels of organization. But, doing so may be difficult.
More specific problems
1. The scaling of ecological processes. Many ecologists include a line about being ‘interested in questions of scale’ on their website blurbs. Despite this, our understanding of the aggregate outcome of multiple processes that are occurring at different spatial or temporal scales remains limited, and poorly predictive. There have been a few useful starts (particularly in Peter Chesson’s scale transition papers (3, 4)), but recent theoretical interest seems to be low. We have data at the community scale, and data at the macro-scale. How do we connect these (and can we)? Models describing how processes occurring at smaller scales produce larger scale dynamics can be complex: they may include non-linearities, autocorrelation between regions, the combination of discrete and continuous events, and multiple attractors.
2. Mechanisms maintaining multi-species coexistence in the real world. Hutchinson’s paradox of the plankton remains unsolved*. Community ecologists have invested a lot of time and energy into understanding species interactions as seen in natural communities. To explore the mechanisms behind coexistence, usually (but not always) ecologists have focused on two-species interactions (or maybe 3): understanding coexistence in larger groups tends to be mostly restricted to theory. But fitting the individual pieces into the larger puzzle is exponentially more difficult: in observed large groups of interacting species, what is the relative contribution of the many coexistence mechanisms identified? Which mechanisms are most important, and how do they change through space and time?
*Perhaps not surprisingly, given it is a paradox...
3. Moving farther away from species. In so many ways, focusing on ‘species’ as the unit of measurement is limiting, because ‘species’ is a discrete term and ecology is interested in quantitative measures. Important advances have been made by redefining ecology as the outcome of species traits and species interactions (5). But I think our ability to connect these ideas more closely to species’ multidimensional niches can still improve. In particular, understanding that traits and interactions can change in context-dependent ways (plasticity, ontogeny, environment) will be important (6, 7).
4. Reproducibility of ecological research. This is more of a philosophical question - how do we achieve reproducibility in a science where context-dependence, alternative stable states, chaos and stochasticity all affect results? How do we differentiate between reproducibility (same results under identical conditions) and generality (same results under similar conditions) in results?
References:
1) Sauer, Uwe; Heinemann, Matthias; Zamboni, Nicola. Genetics: Getting Closer to the Whole Picture. Science 316 (5824): 550–551. doi:10.1126/science.1142502. PMID 17463274.
2) Dominique Gravel, Frédéric Guichard, Michel Loreau and Nicolas Mouquet. Source and sink dynamics in meta-ecosystems. Ecology 91(7): 2172-2184.
3) Chesson, Peter. Scale transition theory with special reference to species coexistence in a variable environment. Journal of biological dynamics 3.2-3 (2009): 149-163.
4) Melbourne, Brett A., and Peter Chesson. The scale transition: scaling up population dynamics with field data. Ecology 87.6 (2006): 1478-1488.
5) McGill, Brian J., et al. Rebuilding community ecology from functional traits. Trends in ecology & evolution 21.4 (2006): 178-185.
6) Poisot, T., Canard, E., Mouillot, D., Mouquet, N., Gravel, D. & Jordan, F. (2012) The dissimilarity of species interaction networks. Ecology letters, 15, 1353–61.
7) Siefert, A., Violle, C., Chalmandrier, L., Albert, C.H., Taudiere, A., Fajardo, A., Aarssen, L.W., Baraloto, C., Carlucci, M.B., Cianciaruso, M.V. and L Dantas, V. A global meta‐analysis of the relative extent of intraspecific trait variation in plant communities. Ecology letters 18.12 (2015): 1406-1419.