There's been some discussion going around ecolog about an
article telling the ecological story of Ascension Island. I should note that the original article is not a great example of science writing; it tries to create conflict that doesn’t exist and lacks a reasonable understanding of ecological theory. There are a couple linked chapters/publications about Ascension Island that make better additions to the story though (
1,
2).
Ascension Island is one of those tiny islands first visited by Europeans in the 1600s. Like many young, small, isolated islands (1200 mi to the next nearest island), it was highly depauperate (~25-30 species of plants). Like many such islands, once humans became regular visitors, new species began to make their to way Ascension. The Brits and their love of cultivating and homogenizing particularly altered the island, and they systematically introduced species calculated to provide ecosystem services, aesthetic value, and food.
As a result, Ascension Island changed strikingly – once an island with lowland deserts and a rocky, barren mountainside, the mountain is today known as Green Mountain. The originally depauperate mountain is now lush with three different vegetation zones, a large variety of plants including “banana, ginger, juniper, raspberry, coffee, ferns, fig trees, Cape Yews, and Norfolk Island pines”, and a complex cloud forest. The original article presents this as some inexplicable outcome, but frankly it seems in keeping with existing ecological ideas. Under island biogeography, if you decrease the distance from an island to the mainland (including via human-aided dispersal), diversity should increase. Given the massive number of species that were introduced, and the coddling they received to aid their establishment, heightened diversity is hardly a surprise. And though the original article suggests that shared evolutionary history is necessary for complex ecosystems, coevolution is hardly a requirement for a functioning ecosystem to develop. Species may be able to coexist despite lacking a shared history--niches may not be filled as tightly as in a long-established, coevolved community, but invasive species research in general should have taught us that novel species combinations can easily occur. Secondly, many of the introduced species on the island are from the same part of the world and likely
do share evolutionary history.
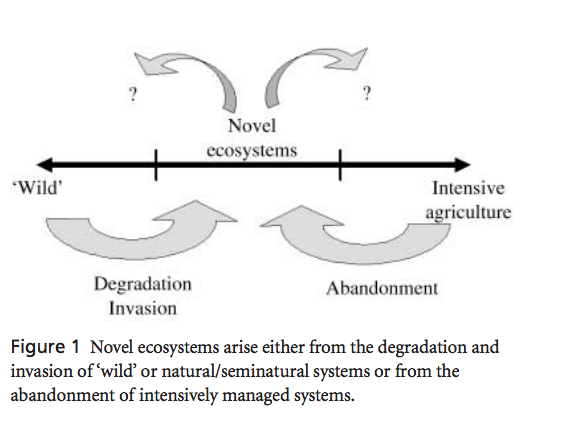
I hadn't given much thought before to the concept of “novel ecosystems” and it has received little attention from the ecological literature (excepting the odd papers, and much more attention from a conservation and management angle). Ascension is a particularly striking example of how human modification leads to ecosystems which are entirely different from anything that has ever been present on the planet. Novel ecosystems have been defined in a number of ways. Generally, they are synthetic ecosystems that include conditions and combinations of organisms never before in existence, and do not depend on human maintenance to persist (as agriculture fields would). Novel ecosystems may be considered to be the outcome of abandonment of human managed systems or else the degradation of existing systems through human activities and invasion (figure). Of course there are incredibly few ecosystems that aren’t affected in some way by human activities (especially in this age of intentional and unintentional human-mediated species introductions), but it is the truly unique ones that are particularly interesting.
There are at least two ways to approach novel ecosystems. One approach is parallel with invasive species and conservation research, and in fact these research areas overlap a fair amount. This is the way in which most research on novel ecosystems seems to be framed. Novel ecosystems carry many of the same issues about making value judgments as invasive species research, and issues of management and whether novel ecosystems can or should be returned to their original state dominate. For example, the conflict between maintaining alpha (island) and gamma (global) diversity exists on Ascension Island– modern, invaded Ascension Island provides greater diversity and ecosystem functioning (erosion control, food, temperature moderation, habitat) than the original barren landscape. But the original endemic species, not surprisingly, have gone extinct or are increasingly at risk.
But focusing solely on these difficult value-laden questions seems to have been at the cost of exploring the value of novel ecosystems as a study system. The most interesting examples of novel ecosystems are not simply modified or invaded ecosystems, but ecosystems that truly never existed before. Like
post-shale dump landscapes in Scotland, where the refuse from mining is now host to unique grasslands that act as refugia for locally rare species; or
the San Francisco Bay, which now is utterly unrecognizable compared to historical descriptions due to heavy invasion; or urban ecosystems with their unique habitats and issues; or even the habitat and connectivity created by
stone fences which now occur on most continents. The questions here aren't always about invasion and management, but instead focus on what the new community looks like. How do novel communities assemble, what processes dominate (mass effects, environmental filtering, competition, predation, etc, etc)? How does ecosystem function relate to the community that assembles? Most BEF research after all, is focused on more traditional ecosystems. What leads to stability in a novel ecosystem, or are they stable at all? They can function is an example of highly unfortunate but also highly informative ‘natural’ experiments for ecologists. But at the moment, if you search for "novel ecosystems" on Google Scholar, the title words are "management", "conservation", "restoration" or "invasion". Actually, there probably are ecologists doing work on novel ecosystems from a purely ecological perspective, but this work gets grouped with disturbance, invasion, and urban ecology: it just remains to consider them in a more unified fashion. If the conversation remains focused only on the conservation issues (as the discussion on ecolog seemed to shift to rapidly), it just seems like we're limiting ourselves a little.