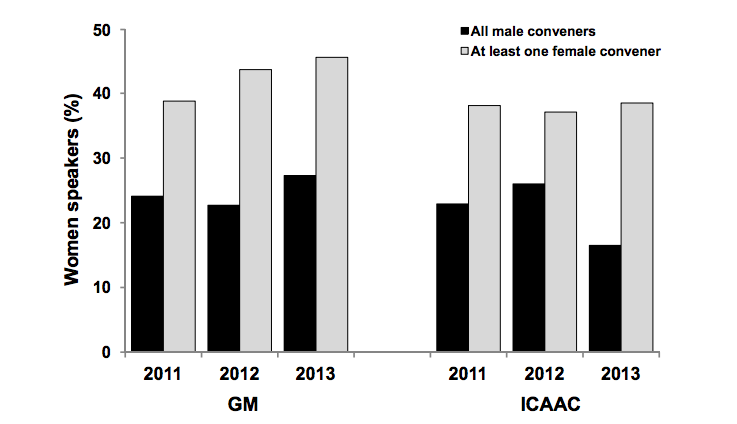
Ecology has been under pressure to mature and progress as a discipline several times in its short life, always in response to looming environmental threats and the perception that ecological knowledge could be of great value. This happened notably in the 1960s, when the call for ecology to be better applicable occurred in relation to the publication of Silent Spring and fears about nuclear power and the Manhattan Project. Voices in academia, government, and the public called for ecology to become a “Big Science”, and focus on bigger scales (the ecosystem) and questions. And yet, “[Silent Spring] brought ecology as a word and concept to the public…A study committee, prodded by the publication of the book, reported to the ESA that their science
was not ready to take on the responsibility being given to it.”
Arguably ecology has grown a lot since then: there have been advances in statistical approaches, spatial and temporal considerations, mechanistic understanding of multiple processes, in the number and type of systems and species studied, and the applications being considered. But it is once again facing a call (one that frankly has been ongoing for a number of years) to quickly progress as a science. The Anthropocene has proven an age of extinctions, human-mediated environmental changes, and threats to species and ecosystems from warming, habitat loss and fragmentation, extinctions, and invasions abound. Never has (applied) ecology appeared more relevant as a discipline to the general public and government. This is reflected in the increasing inclusion of buzzwords like “climate change”, “restoration”, “ecosystem services”, “biodiversity hotspot”, or “invasion” as keys to successful self-justification. Also similar to the 1960s is the feeling that ecology is not ready or able to meet the demand. Worse, that the time ecology has to respond is more limited than ever.
This first point--that ecology isn’t ready--is repeated in Georgina Mace’s (the outgoing president of the British Ecological Society)
must-read editorial in Nature. The globe is in trouble, from climate change, disease, overpopulation, loss of habitat and biodiversity and Mace argues that ecology is incapable in its current form of responding to the need. She suggests that unless ecology evolves, it will fail as a discipline. Despite the growth of ecology that followed the 1960s, it is still a 'small' discipline: collaborations are mostly intra-disciplinary, data has been privately controlled, and the tendency remains to specialize on a particular system or organism of interest. However, this 'small' approach provides very little insight into the big problems of today - particularly understanding and predicting how the effects of global change on ecosystems and multispecies assemblages. To Mace, the solution, the undeniable necessity, is for ecology to get bigger. In particular, collaborations need to be broader and larger, with data sharing and availability (“big data”) the default. Ecological models and experiments/observations should be scaled up so that we can understand ecosystem effects and identify general trends
across species or systems. In this new 'big' ecology, “[g]oals would be shaped by scientists, policy-makers and users of the resulting science, rather than by recent publishing trends”. Making research more interdisciplinary and including end-product users would allow the most important questions to receive the attention they deserve.
The difficulty with the looming environmental crises and the pressure on ecology to grow, is that the important decisions to be made have to be made rapidly and perhaps without complete information. Often scientific progress is afforded the time for slow progression and self-correction. After all, change is costly and risky, it requires reinvesting effort and funding, and may or may not pay off, and so science (including ecology) is often conservative. For example, a conservative mind would note that Mace’s suggestions are not without uncertainty and risk. Big data, for example,
is acknowledged to have its strengths and its weaknesses, it may or may not be the cure-all it is touted as. Regardless of the amounts of data, good questions need to be asked and data, no matter how high quality, may not be appropriate for some questions. Context is often so important in ecology that attempts to combine
data for meta-analysis may be questionable. Long running arguments within ecology reflect the fear that making ecological research more useful for applications and interdisciplinary questions may come at the expense of basic research and
theory. It seems then that ecology is in an even worse scenario than Mace suggests, since not only must ecology change in order to respond to need, but it also must predict with incomplete information which future path will be most effective.
So ecological science is at an important junction with choices to make about future directions, limits on the information with which to make those choices, little time to make them, and much pressure to make them correctly. Perhaps we can take some comfort from the fact that ecology has been here before, though. There are some lessons we can draw from ecology’s last identity crisis, both the successes and failures. The last round resulted in ecology gaining legitimacy as a science and being integrated into policy and governance (the EPA, environmental assessments, etc). It appears, particularly in some countries, that ecology is more difficult to sell to policy and government today, but at the very least ecology has established a toehold it can take advantage of. Ecology also tried to focus on bigger scales in the 1960s--the concept of the '
ecosystem' resulted from that time--but the criticism was that the new ideas about ecosystems and evolutionary ecology weren't well integrated into ecological applications, and so their effect wasn't as broad as it could have been. Concepts like ecosystem services and function today integrate ecosystem science into applied outputs, and the cautionary tale is the value of balancing theoretical and applied development. It also seems that ecology must first consider what its duty as a science is to society (Mace’s assumption being that we have a great duty to be of value), since that is the key determinate of what path we decide to take. Then, we can hopefully consider what have we done right in recent years, what have we done wrong, and then decide where to go from here.
 |
| Page from "Silent Spring", Rachel Carson. |